
「塩鶴」
なぜ、鶴を塩漬けに???
文字通り、鶴を塩漬けにして食用にしたものです。
食べるものがなかったからといって、鶴を食べることはないじゃないかと思う人は多いのではないでしょうか。
しかし現在でも賛否ある中でクジラの商業捕鯨が30年ぶりに解禁になったり、他国では犬を食用にしたり、一方ではあらゆる肉を食することを禁止したり、古今東西でさまざまな食文化が見られます。
それぞれ文化的、思想的背景があって食用にしていたはずですが、では鶴はどのような背景で食用にされていたのでしょうか。
タンチョウヅルと言えば、現在では北国の風物詩、冬の訪れを告げる象徴的な生き物とされています。
しかし、かつてタンチョウは日本各地に広く分布していました。これを証明するように、江戸時代の資料では東西を問わず日本各地でタンチョウに関する記録を確認することができます。
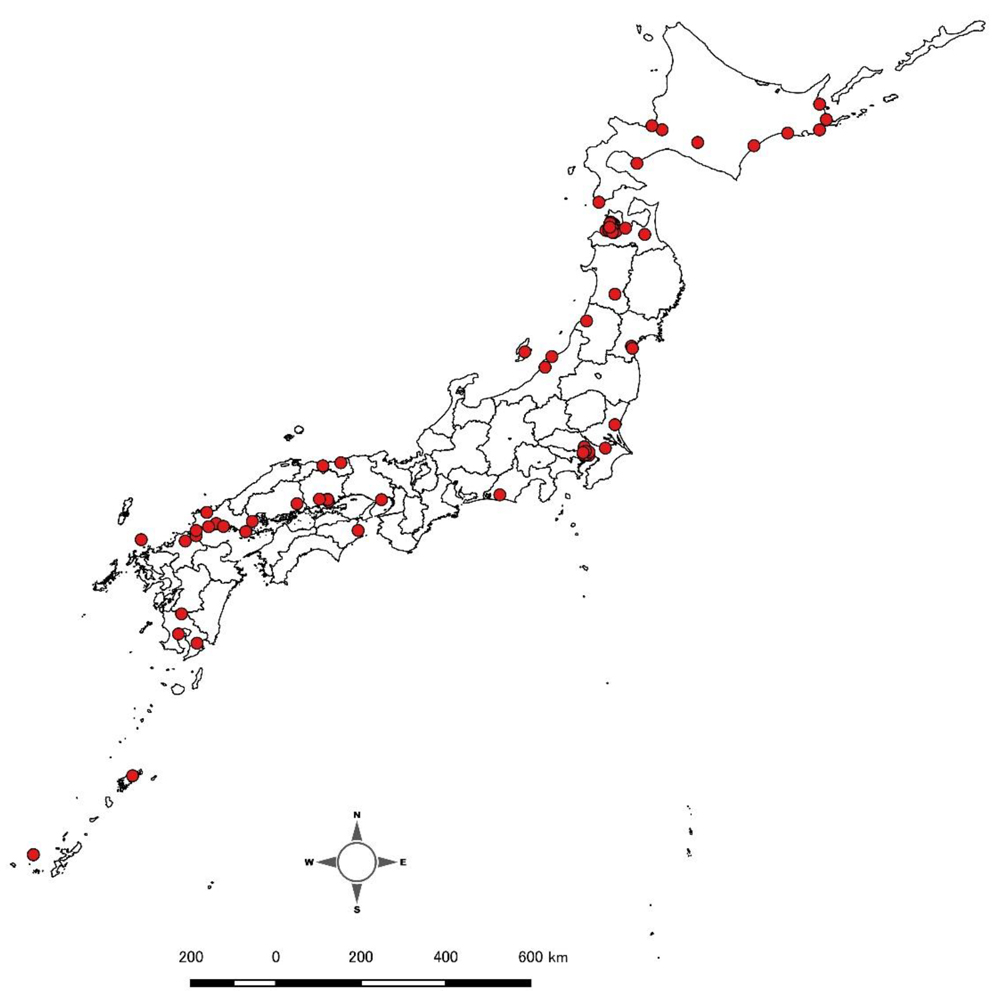
図9 江戸時代におけるタンチョウの分布復元図。江戸時代の文献資料からタンチョウに関する記録が得られた地域を●で示した。なお、現状では●がない地域についても、今後情報が追加される可能性があり、過去の分布を否定するものではない。
たとえば、台東区の三ノ輪でもその姿が見られたようで、歌川広重の筆による「箕輪金杉三河しま」には、特有の紺青の色彩とともに描かれています。

図7.『名所江戸百景』より「箕輪金杉三河しま」。タンチョウが渡来する豊かな自然環境が江戸周辺にも残っていたことを説明する際に用いられることが多いが、江戸周辺のツルは人為的な管理下にあった可能性も否定できない。
国立国会図書館デジタルコレクションより
ただこの地域などで見られたタンチョウヅルは、野生のものかは定かでなく、将軍の鷹狩のために餌付けされていた可能性が高かったようです。
鶴を食べる
このように人間の手によって管理されていたタンチョウヅルですが、この種類自体が食されたことは珍しかったようです。
というのも、鶴の中にも美味とそうでないものがあり、最も好まれたのはナベヅル。
タンチョウヅルの場合は、
「丹頂の者は肉硬く、味い美ならず」
とされている通り、肉は硬く、美味しくなかったようです。
タンチョウヅルはむしろ、籠の中、池、庭で鑑賞されることが主な用途だったようです。
歴史鳥類学という領域では、歴史資料から当時の鳥類の生態を推測し、同時に人と鳥との営みを歴史も垣間見せてくれます。
希少な鳥類の保全が叫ばれて久しいですが、この領域による知見からはどのような経緯で今に至ったのかを知らせてくれます。
