
ここでは本の発売に先駆けて、「第4章 閉ざされた湖で起こった進化」をご覧になれます。ぜひご覧いただいて、本書を手に取る参考にしてください。
前章まで、サメ、真骨魚、古代魚の進化など、1000万年から1億年単位で起こってきた大きな進化について紹介してきた。本章では、ペットショップでもおなじみで、世界中のアクアリストから観賞魚として寵愛されている「シクリッド」の進化をモデルに、短期間(といっても数万年)で急激に起こった小さな進化について紹介する。
シクリッドとはスズキ目カワスズメ科に属する魚類の総称で、南米からアフリカ、マダガスカル、東南アジアまで旧世界の広くにその分布域を広げている。このシクリッドにはエンゼルフィッシュやディスカスなど一般にも多くの人がよく知る熱帯魚も含まれているが、筆者のような進化研究者が特に興味をもっているのが東アフリカ産シクリッドである。東アフリカの三大湖(ビクトリア湖、マラウィ湖、タンガニィカ湖)には数百をこえる固有のシクリッドが生息しており、種の分化や多様化が驚異的なスピードで起きたことが知られている。つまりシクリッドは進化のメカニズムを理解する上で理想的なモデル生物といえよう。一見するとモヤモヤとしてつかみどころのない「進化」という現象をいかにしてDNAレベルで理解していくのかを、シクリッド進化における著名な研究者たちの論文を紹介しながら以下に解説していく。まず、進化の舞台でもある東アフリカ三大湖の形成の歴史を紹介する。どのような地理的環境がシクリッドの驚くべき多様化を可能にしたのであろうか?
1.シクリッドの多様性創出とその要因
⑴シクリッド進化の舞台―東アフリカ三大湖の形成と地理的な歴史
シクリッド多様性創出の場となった東アフリカ三大湖は、その大きさで世界有数の規模を誇っているだけでなく、湖ごとにその環境や成立年代が大きく異なっていることが東アフリカ産シクリッドの多様性に大きな影響を及ぼしていると考えられる。東アフリカの三大湖は、アフリカ大陸を南北に縦断する大地溝帯(グレートリフトバレー)の形成と深く関わっており、大陸が分裂するように働く力によって形成された深い裂け目に水が流入してできたのが、タンガニィカ湖やマラウィ湖であり、これらはいわゆる「地溝湖」と呼ばれている。それに対してビクトリア湖は、東西の地溝帯に挟まれた場所に位置しており、両地溝帯の形成に伴って発生した隆起が陥没することで誕生した湖であると考えられている(図1A)。そのため、ビクトリア湖は水深も80m程度であり、他の湖(タンガニィカ湖、マラウィ湖はそれぞれ570m、290m)と比べると浅いため、湖の透明度も低く水位の変動による湖環境への影響が大きいことが知られている。各湖の成立年代に関してはこれまでに多くの地質学的研究がなされており、最も古いタンガニィカ湖の成立が今から1200万〜900万年前と推定されている。そしてマラウィ湖は450万年前にはすでに成立していたものの、160万年前には一度完全に干上がりそこからまた水が流入することで再び湖が形成されたと推定されている。ビクトリア湖の成立年代はさらに新しく、今から1万2400年前に完全に干上がっていた盆地に水が流入してできたという報告があり、現在は大きな岩石の堆積した独特な生態的環境をもっている(図1B)。つまり、三大湖の成立年代はそれぞれが大きく異なっており、また現在は湖どうしが河川でつながっていることはなく、湖間での生物相の流入は起きていないため、各湖に生息する数百種ものシクリッドは全て固有種となっている。後述するが、以上にあるような三大湖の地質学的な特殊性や成立年代の違いが、シクリッドの系統進化パターンとも深く関わっていることがわかり始めている。
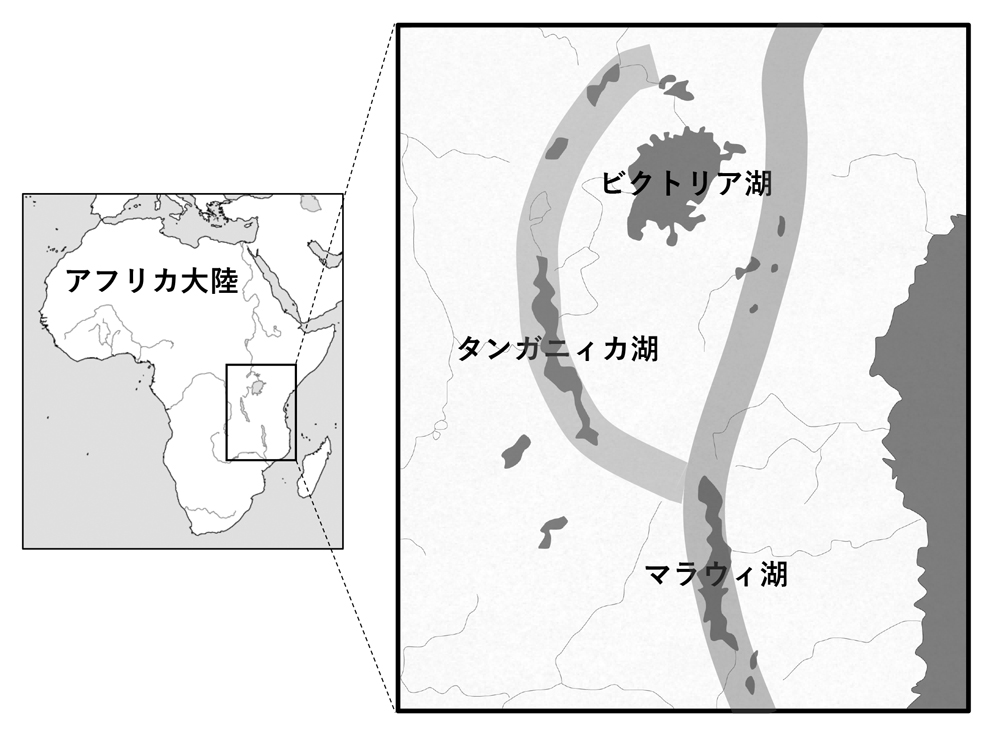
図1A.東アフリカ三大湖と大地溝帯。
タンガニィカ湖とマラウィ湖は大地溝帯の裂け目に存在する地溝湖。それに対してビクトリア湖は隆起地帯の陥没によって生じた湖。灰色太線は大地溝帯を表している。

図1B.ビクトリア湖の様子。巨大な岩が積み重なっているのが特徴で、他湖と比べて透明度は低い。ロックシクリッドと呼ばれる岩場種が多数生息している。
⑵シクリッドの美しい婚姻色とその進化のメカニズム
本章のメインともいえるシクリッドの多様化メカニズムを紹介するにあたり、まずシクリッドの生態的多様性そのものをいくつかの例に絞って以下に紹介する。ここに挙げるほんの数例を概観するだけでも、東アフリカ産シクリッドが、どれだけの多様性を誇っているのかを容易に感じ取ることができるだろう。また、このシクリッドの多様化は様々な湖環境に適応して生み出された自然選択による産物なのであることはいうまでもない。
シクリッドにおいて観察される多様な特徴の中でも、研究者たちが最も注目してきたのがオスの婚姻色であろう。シクリッドは一般にオスが性成熟に達すると派手な婚姻色を呈するのに対し、メスは性成熟にいたっても隠蔽的な体色しかもたず、そこには明らかな性的二型が観察される(図2)。この性的二型は、オスが繁殖の際に婚姻色を呈してメスにディスプレイし、メスはオスの婚姻色を選択して交配するという、選択的交配によって生み出されたと考えられる。また、オスの婚姻色が種によって多様であることや、婚姻色のバリエーションが多いシクリッド系統は種数も多いことから、メスによるオスの婚姻色の選択、つまりは性選択がシクリッドの種分化をメインに駆動してきた可能性が示唆されている。婚姻色とそれを認知する視覚、そして性選択に関わる諸問題は種分化を始めとする進化現象の多くを理解する上で重要な形質であるため、それらに関するDNAレベルでの研究については後ほどあらためて紹介していきたい。

図2.シクリッドの婚姻色。オス(上段)はそれぞれの種に特徴的な婚姻色を呈する一方、メス(下段)は地味な隠蔽色をもつ。また、尻びれのエッグダミーについてもオスにおいて特に目立つ配色となっている。図に示したのはビクトリア湖シクリッドで、左からLithochromis rufus, Haplochromis pyrrhocephalus, H. chilotes。
⑶シクリッドに特徴的な交尾行動と口内での哺育
シクリッドにおいてよく知られている行動に口内哺育がある。一般的に魚類は水草等に卵を産み付ける基質産卵を行い、程度の差こそあれ孵化後の稚魚は自由に水中を泳いで成長していくことが知られている。しかし、東アフリカ産シクリッドの多くは(ビクトリア湖、マラウィ湖では実に100%、タンガニィカ湖では40%)、メスが受精卵をみずから咥え稚魚が十分に育つまで口の中で哺育をする「マウスブルーダー」である。これは、卵や稚魚を捕食から防ぐという点から考えて極めて適応的な子育て戦略であると考えられる。この口内哺育を達成するためには、母親の口の中で精子と卵が受精するイベントが起きなくてはならないのだが、それを可能にしているのがマウスブルーダーに特徴的な交尾様式であり、その一連の流れを図3に示す。まず交配相手の決まった雄雌のシクリッドは共に尾部を激しく振動させながらお互いの総排出腔を相手の鼻先に提示し、クルクルと円を描くように回転する。やがてメスの総排出腔から一度に数粒の卵が産み出されるわけであるが、メスシクリッドは回転の最中にこの卵を拾い上げて口に咥える。このメスが卵を拾い上げる際に、オスは尾部を振動させながら尻びれにある円形のスポット模様(エッグダミー)をメスにアピールする。するとメスはこのエッグダミーをみずから産んだ卵と勘違いし、それを咥えようとして口を近づける。その際にオスは総排出腔から精子を振りまくことで、メスの口の中で受精が完了するという、なんとも絶妙なタイミングで口内哺育という特異な繁殖システムが完了するわけである。この回転しながらの特徴的な交尾は10分程度続けられ、最終的には数十個ほどの卵が産み出され、それらは母親の口内で育て上げられることになる。
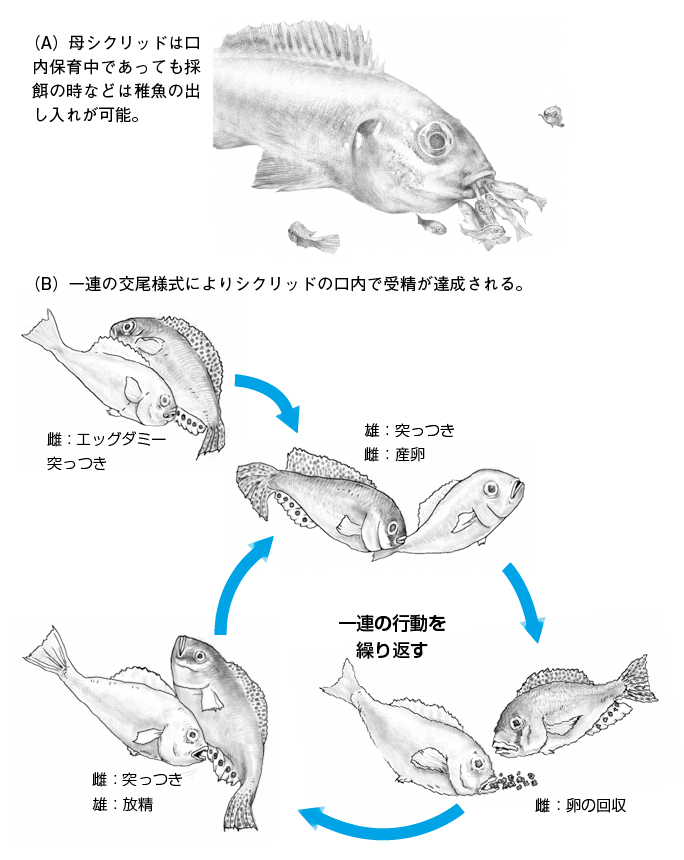
図3.シクリッドに特徴的な口内保育とそれを可能にする交尾様式
このマウスブルーディングは進化的にも発達した哺育行動であると考えられるが、このシクリッドの哺育行動を逆手にとって利用する種がいる。これは佐藤哲博士によって発見されNatureに発表されたナマズの托卵行動である(1)。ナマズの一種であるSynodontis multipunctatusは、マウスブルーダーの産卵現場に突入し、シクリッドの卵の中に自分の卵を紛れ込ませる。母親シクリッドの口内に潜り込んだナマズ卵の発生は、本来の子であるはずのシクリッドのものより数段早く進み、成長に伴ってナマズがシクリッドの稚魚を捕食してしまう。最終的に母親シクリッドは大きく成長したナマズの子のみを育てることになるのだ(図4)。明らかに外見の異なるナマズの稚魚をなぜ母親が識別し排除し得ないのかは、現在のところよくわかっていないが、シクリッドにおいて巧みに進化した口内哺育行動をさらに利用したナマズの托卵行動という、まさにシクリッドの世界の多様な一面である。

図4.ナマズの一種によるシクリッドへの托卵。(上)托卵ナマズSynodontis multipunctatus 提供:高橋鉄美博士(兵庫県立大学)。(下)メスシクリッドの口内から見つかったSynodontisの稚魚。提供:宗原弘幸博士(北海道大学)
⑷親以外が参加する子育てシステム―シクリッドの協同繁殖
もうひとつ、シクリッドの多様な生態を紹介する上で欠かせないのが協同繁殖であろう。協同繁殖とは、親以外の個体が世話をする行動のことで、いわゆる利他的な行動の進化を理解する上で重要である。この協同繁殖は、アリやミツバチ、シロアリなどの真社会性昆虫においてよく知られた行動で、脊椎動物では主に哺乳類(ハダカデバネズミ、ミーアキャット)や鳥類で研究が進んでいるものの、魚類ではあまり知られていなかった。その中で、1981年にTaborskyとLimbergerによってタンガニィカ湖に生息するシクリッドNeolamprologous brichardiの繁殖にはヘルパーが存在していることが明らかにされた(2)。もちろん魚類におけるヘルパーの存在はこれが初めての報告であり、その後は他のシクリッドにおいても協同繁殖が確認されている。
協同繁殖を行うシクリッドはひとつのなわばりを共有する群れ(家族)を作るが、この群れには子育てをする大型の個体に加えて、なわばりに侵入してきた異種他個体や他の群れ個体を追い出す小型の個体、つまりはヘルパーが存在していることがわかった。協同繁殖については、その個体自身ではなく、遺伝子を共有する血縁個体の繁殖成功を増すといういわゆる「血縁淘汰」によってその多くを説明できると考えられているが、実際にシクリッドの群れにおける血縁度をDNAレベルで観察した研究では、ヘルパーの年齢が増すにつれて繁殖親との血縁度が下がることも明らかとなり、その繁殖様式は予想以上に複雑であると考えられるようになってきた。さらには、ヘルパーのシクリッドは営巣地を取り囲む石の下に、無脊椎動物が集まるような穴を整備することで群れ全体の餌資源を増大させるという高度な社会性を獲得している種もいることがわかった。脊椎動物の進化を考える上で、哺乳類や鳥類よりもむしろ原始的なグループ(語弊があるかもしれないが、あえて原始的という言葉を選んだ)における社会性を研究することで、その遺伝的なメカニズムを起源や進化にまで深く掘り下げながら理解することができることから、このシクリッドの共同繁殖の研究が今後も精力的に進んでいくことを期待したい。
⑸シクリッドにおいて進化した驚くべき食性や歯の多様性
東アフリカの各湖のシクリッドはお互いに近縁なグループであるが、その遺伝的な近縁性とは裏腹に食性においては著しい多様性が観察される。さらに、食性の多様化に伴って歯や顎、さらには咽頭歯(一般的な歯に加えて喉にある歯でコイやベラの他にシクリッドで発達している)の形態も多様化を遂げていることは極めて興味深い。その一例を図5に示す。シクリッドは大きくプランクトン食、魚食、藻類食、貝食などが挙げられるが、例えば魚食性のシクリッドは、体長と比して吻部が大きく体も長い種が多い。藻類食の種は岩の表面に生えたコケ等を削り取って食べるために、歯列は細かく密であり、吻部そのものが下側を向いている。また貝食シクリッドは、硬い殻を割るために咽頭歯は他種と比較して太く頑丈な構造に進化している。食性の多様化は、餌資源をめぐる種間競争を緩和することから、複数種の共存を可能にすると考えられ、シクリッドの生息する湖という限られた環境下において種多様性を生み出す機構として重要な意味合いをもつのは間違いない。
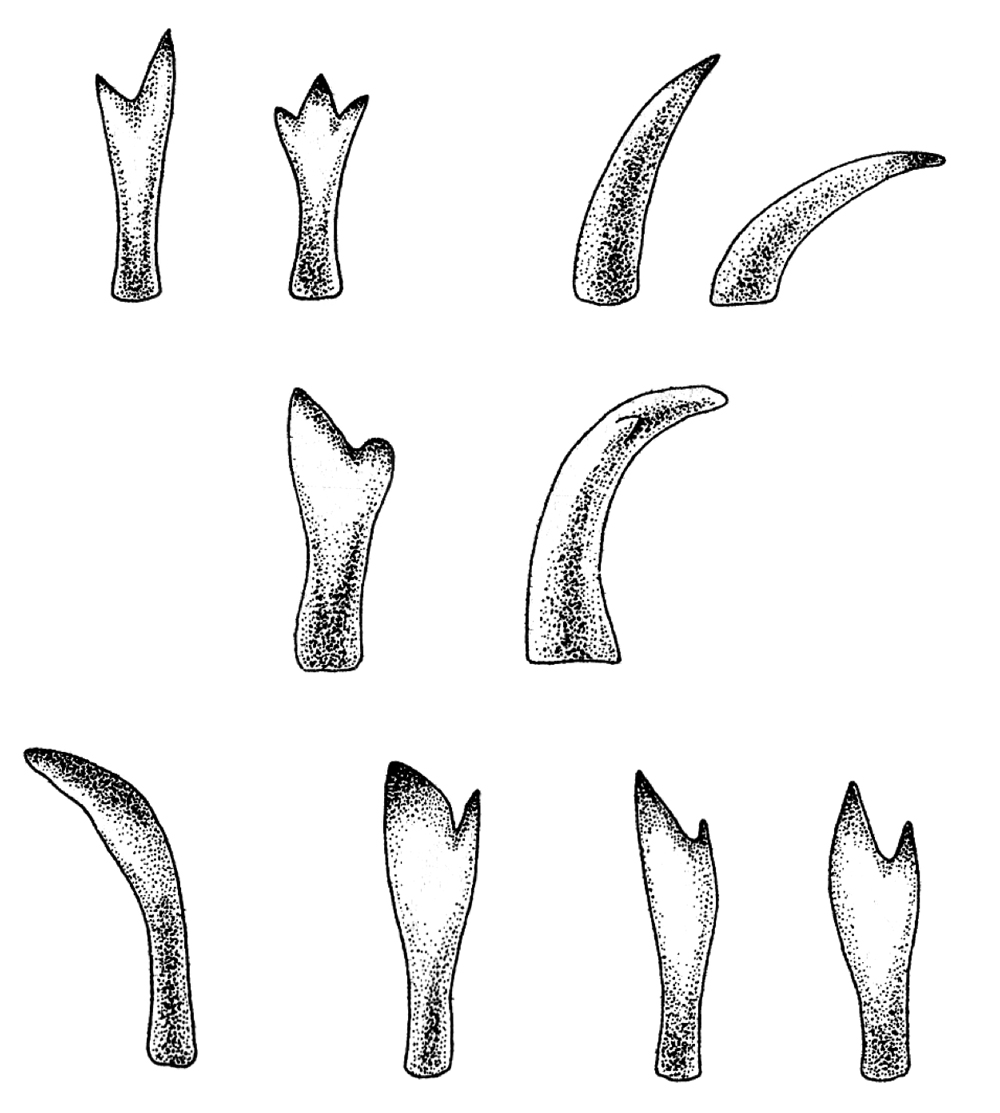
図5.ビクトリア湖シクリッドにおける多様な歯式の例
そして上記のようなシクリッドの多様な食性の中でも特に研究者の興味をひくのが、タンガニィカ湖のPerisodus microlepisに代表されるシクリッドで観察される鱗食いである。この魚種は他の魚のウロコをはぎ取って食べるという極めて特殊な生態をもつ。さらに興味深いことに、このシクリッドは鱗を食べる際の攻撃の方向に右利きと左利きの個体が存在しており、それが遺伝的な支配を受けているという。京都大学の堀道夫博士の長年の調査の結果、P. microlepis種内にはこの右利きと左利きが常に存在し、その割合が周期変動することがわかった(3)。これは、右利きの個体数が増えた場合に、捕食される側が学習することで右利きの捕食成功率が下がり、今度は相対的に有利となった左利きの個体数が増えるからと考えられる。右利きと左利きの個体数に周期的な変動が生じる。これは少数個体が有利となる負の頻度依存淘汰として知られている。これまで、堀博士により発見されたシクリッドの利き口は多くの研究者が興味をもって研究を続けてきたが、そのDNAレベルでのメカニズムは現在にいたっても明らかにされておらず、今後の研究の進展が期待されている。
2.DNAに刻まれたシクリッドの多様性の「歴史」
⑴シクリッドの進化の歴史を紐解く分子系統学
これまでは東アフリカ産シクリッドの驚くべき多様性を生態や行動レベルで紹介してきたが、これからはその多様性の遺伝的基盤を探るべく進められたDNAレベルでの研究を紹介する。進化学に分子生物学的手法が応用され始めて30年近くが経とうとしているが、シクリッドのDNA研究ではいつの時代も常にエキサイティングな発見が続いている。
シクリッドのDNA研究の歴史はその分岐順序、つまりは系統関係を解き明かすところから開始された。最も初期の論文はMeyerら(4)によるビクトリア湖シクリッドの起源に関する研究である。上述した通り、ビクトリア湖は他の湖と比べて地質的な歴史が新しく、そこに生息する多様なシクリッドがどうやって誕生したのか、つまりは単一の起源なのか、それとも他湖のシクリッドが複数回移入してきたものなのか、といった系統学的な問題に興味が集まっていたが、その詳細はほとんど明らかにされていなかった。タンガニィカ湖に生息する一部のシクリッドとビクトリア湖やマラウィ湖のシクリッドはHaplochromineと呼ばれる大きなグループに分類され、その種数は地球上の全ての真骨魚類の7%にも達する極めて繁栄したグループである。
その当時の形態学的な研究によれば、ビクトリア湖のシクリッドは起源が単一ではなく、湖周辺に生息している多様化したHaplochromineシクリッドが複数回移入して形成された多系統群であろうと考えるのが一般的であった。しかし、Meyerら(4)によって進められたミトコンドリア部分配列の分子系統解析の結果、形態的に極めて多様なビクトリア湖のシクリッドは単系統群(祖先種に由来する全ての種をメンバーとして含むグループ)を形成することが強く示唆され、つまりはその多様性が湖の成立後の極めて短期間に獲得されたものであるという驚くべき仮説が提唱された。その後も東アフリカ三大湖のシクリッドに関する分子系統解析は続けられ、その系統的な進化史については概ね一致する結果が得られるようになり、最終的にはSalzburgerら(5)が提唱したOut of Tanganyika仮説に集約された。その仮説によれば、東アフリカ三大湖の中で最も繁栄したHaplochromineシクリッドは、最も古いタンガニィカ湖にその起源をもっており、タンガニィカ湖の水位が上昇していた時期(350万〜110万年前)に、その祖先となるグループが河川を通じて湖から溢れ出すかたちで生息地を拡大し、現在ではその子孫となる種がビクトリア湖やマラウィ湖、とその周辺河川に分布するにいたったということである。そして、現在において最も代表的な系統樹(図6)によれば、「Out of Tanganyika」を遂げたシクリッドはマラウィ湖、ビクトリア湖で独立に放散を遂げたため、それぞれが単系統群を形成する。特にビクトリア湖のシクリッドは、その周辺に存在し、小さいながらも古くから存在するキブ湖に起源をもつことが示された(6)(図7)。また、一度タンガニィカ湖から流出したグループが再度タンガニィカ湖に戻ったケースもあったようで、それが現在ではTropheiniと呼ばれひとつの族(Tribe)を形成している。ちなみに「Out of Tanganyika」は人類史における「Out of Africa」に習ったものである。Salzburgerらは河川を通じてOut of Africaを達成したシクリッド祖先種において獲得されたエッグダミーや口内哺育などの鍵革新的な形質が生態的機会と協調的に働いて、その後の湖環境における適応放散につながったと考察している。
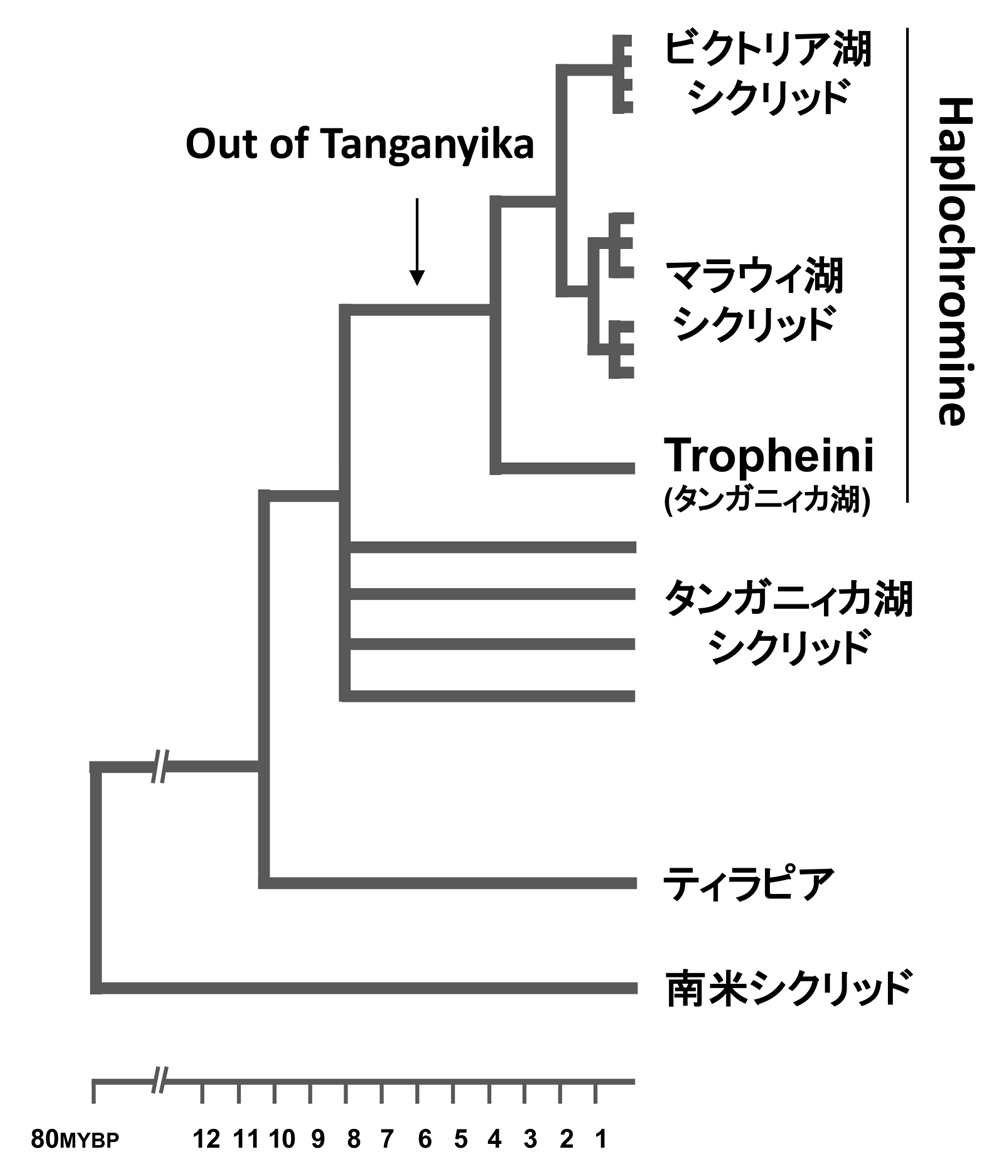
図6.東アフリカ産シクリッドの系統樹(文献10を改変)
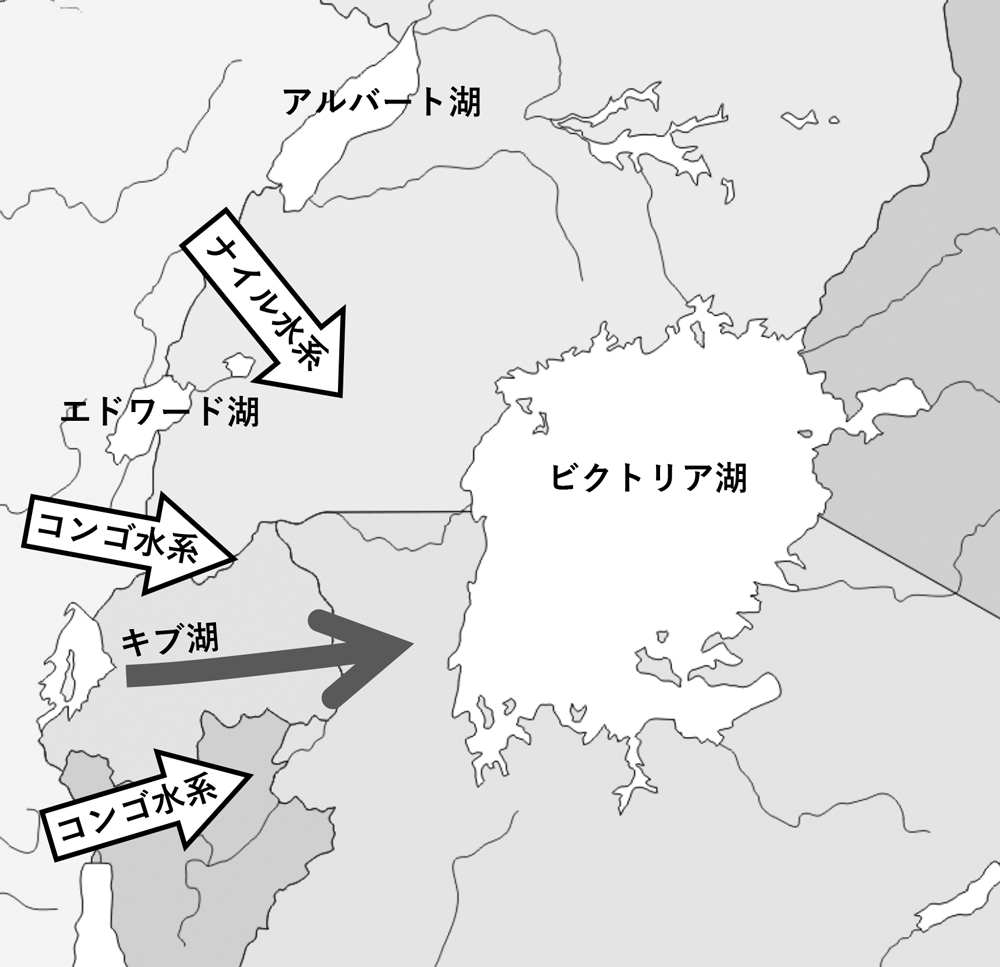
図7.ビクトリア湖シクリッドの起源に関する対立仮説Verheyenらのキブ湖単一起源仮説(灰色矢印)と、Seehausenらのコンゴ・ナイル水系の交雑起源仮説(白矢印)(文献11と13より)
東アフリカ三大湖シクリッドの系統進化に関する上記の仮説は多くの研究者においてコンセンサスが得られているが、唯一ビクトリア湖の起源についてはSeehausenのグループによる核ゲノム解析によって異なる仮説が提唱された(7)。シクリッドの分子系統学的解析の多くは、組み換えがなくハプロイドであるために系統樹推定において扱いやすいミトコンドリアのDNA配列を指標として進められてきたが、ビクトリア湖シクリッドのように比較的最近に複雑な進化過程を経たグループについては、単純化され過ぎたストーリーを導き易い。Seehausenのグループは核ゲノムを対象としたAFLP解析により、ビクトリア湖やマラウィ湖のシクリッドの系統推定を行ったところ、最も予想外であったのが、ビクトリア湖シクリッドの遺伝的多様度が高いことで、その多様度はマラウィ湖シクリッドと同程度であった。ミトコンドリア配列に基づく解析によれば、ビクトリア湖シクリッドの多様性は極めて低く、それはビクトリア湖が新しいという地質学的とも一致していたのだが、核ゲノムの解析結果はそれと大きく食い違う結果を提示したことになる。また系統樹上においては、コンゴ水系とナイル水系の河川に生息し生態的にも多様なThoracochromis属のシクリッドがビクトリア湖シクリッドの姉妹群に位置していた。この系統樹からSeehausenらは、ビクトリア湖シクリッドの起源がナイルとコンゴ水系の河川に生息していたThoracochromis属のふたつの系統が交雑することよって生じたという「交雑起源仮説」を唱え(図7)(5)、Salzburger(6)やVerheyenの唱えたキブ湖単一起源説を完全に否定した。その後もSeehausenのグループは全ゲノムレベルでこの問題に取り組み、やはりビクトリア湖シクリッドの交雑起源説を示唆する証拠を得ている(8)。
東アフリカ三大湖シクリッドの系統地理学的な流れとしては、Out of Tanganyikaによりシクリッドの生息域拡大があり、そこからビクトリア湖に流れ込むタイミングで大規模な交雑が起きたということで説明がつくのではないかと筆者は考えている。ただし、ナイルとコンゴ水系にいる2グループの交雑を可能にしたゲノム基盤や、マラウィ湖の起源についても同じように説明が可能なのかについても、まだ議論する余地は多分に残されており、今後のさらなるサンプル採集や全ゲノム配列に基づいた網羅的な解析に期待するところである。
⑵失われた古代湖Makgadikgadiの再発見とそこで起きた適応放散
もうひとつ、東アフリカでなく南アフリカの河川に生息するシクリッドに関して、系統地理学的に興味深い話題を紹介したい。東アフリカの河川に生息するシクリッドは湖のシクリッドと比較して形態的・生態的な多様性が低く、これは湖と河川における生態学的な機会の差を如実に反映しているものと考えられてきた。しかし、不思議なことに南アフリカの河川には東アフリカの湖と同程度に多様なシクリッドが生息していることが知られており、これはいかなる理由であるのかがわかっていなかった。
そこでSeehausenのグループはミトコンドリア配列を用いて、東アフリカおよび南アフリカのシクリッドの系統解析を行ったところ、南アフリカの多様なシクリッドが単系統群を形成することが明らかとなった。この結果を受けJoyceら(9)は、現在は河川という地理的に離れた場所に生息している南アフリカのシクリッドは、実は完新世(約1万年前)に干上がったと考えられている古代湖Makgadikgadiで放散を遂げた系統であるとの仮説を提唱した。現在、ボツワナのカラハリ砂漠に位置するMakgadikgadiは乾燥した塩湖となっているためシクリッドが生息することはできないが、かつてそこに存在した古代湖において適応放散を遂げたシクリッドが、湖は干上がってしまった現在においても多様性をそのまま保ちながら周辺河川で生き延びているのだと結論づけた。同時に、湖という環境がもつ生態学的機会がシクリッドの多様化に及ぼす影響は極めて大きいことを強調した。アフリカ大陸は地殻活動が盛んであり、巨大な湖の出現や消失などダイナミックなレベルの環境変動が激しいため、現在だけを観察しているだけでは気づくことのなかった進化ストーリーが分子系統樹を通して垣間見えてくるのもアフリカ産シクリッド研究の楽しいところである。
⑶シクリッドの分子系統樹から明らかになった「平行進化」
これまで紹介してきたように、DNAを用いた系統樹推定から、東アフリカのシクリッドが各湖で適応放散を起こしたことが明らかにされたが、もうひとつの大きな発見は驚くべき平行進化が起きていたことである。Kocherら(10)はタンガニィカ湖、マラウィ湖には形態的に似通ったシクリッドが存在していることに着目し、それら形態の似たシクリッドどうしが遺伝的に近縁なのか、それとも似た形態はそれぞれの湖で独立に進化したものなのかを検証した。分子系統樹については詳細を上述した通りで、結論から述べると後者の仮説が正しかった。つまり、マラウィ湖に生息しているシクリッドは単系統群を形成することから、驚くほど似通ったシクリッドはそれぞれがタンガニィカ湖とマラウィ湖において独立に進化したものであり、まさに典型的な平行進化(異なる種同士が独立に似通った方向へ進化を遂げること)が起きていることが示された。これについてはビクトリア湖にも当てはまり、タンガニィカ湖、マラウィ湖、ビクトリア湖では、あらゆる形質が平行進化(歯列、顎角度、唇の厚さ)したものであることが明らかになっている(図8)。それぞれの湖における似た環境が提示する生態学的機会が、ここまで如実な平行進化を駆動していくという事実は極めて興味深い。ただし、この平行進化のDNAレベルでのメカニズムについても明らかにされていることは少なく、現在でも筆者らを含めた多くの研究者がこの問題に取り組んでいるところである。
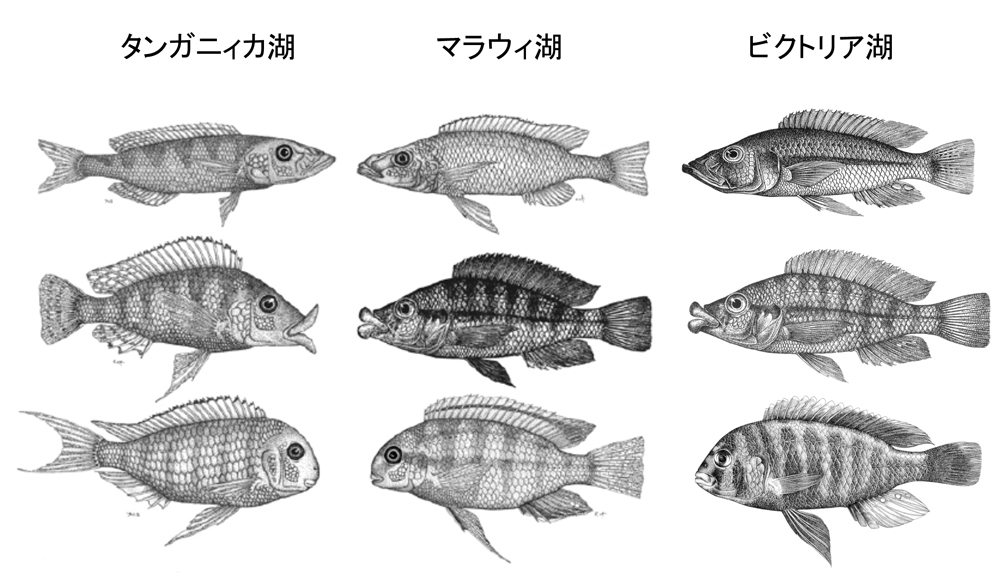
図8.東アフリカ三大湖シクリッドで観察される平行進化。湖間で驚くほど似た形態のシクリッドが観察されるが、遺伝的には各湖のシクリッドどうしが近縁である。
3.DNAに刻まれたシクリッドの多様性の「メカニズム」
⑴カモフラージュ遺伝子と性との関わり合い
分子系統学によってシクリッドの系統地理学的な問題、つまりどのような順序で多様性が生み出されてきたのかについてはその多くが明らかにされ、適応放散と平行進化というふたつの注目すべき現象が起きているということがわかった。この分子系統学的な多くの発見に続く次なる興味は、どのようなメカニズム(遺伝子の変異や発現パターンの変化など)によって形態的な多様性が獲得されたのかというところだ。これは進化発生(Evo-devo)と呼ばれる学問分野であり、シクリッドの分子系統推定を行ってきた研究者の多くが2000年頃を皮切りにこのエボデボ研究にも参戦を始めた。

図9.主にメスのシクリッドで観察される斑模様(OB morph)。上がオスの野生個体、下がメスのOB個体。ともにマラウィ湖シクリッドの1種Maylandia tarakiki。
シクリッドにはオレンジブロッチ(OB)と呼ばれる奇妙な模様をもった個体が主にメスにおいて一定の頻度で出現することが知られている(図9)。このOB模様をもった個体は色合い的に生息域にある岩と同化するため、捕食者からのカモフラージュとしての役割があると予想されている。しかし、シクリッドのオスは、その婚姻色が生殖相手の獲得に最も重要な因子のひとつであるため、オス個体にこのOBが出現することは望ましくない。そのため、メス個体のみでこのOBが出現するようになったと考えられ、いわゆる性的対立をどう解消しているかという進化学的にも興味が集まっている現象である。しかし、2000年初頭の頃にはその遺伝的なメカニズムは当然のことながら明らかにされていなかった。この問題に取り組んだのがStreelmanらであり、彼らはまずマラウィシクリッドの通常オスとOBメスF0世代を掛け合わせ、その孫(F2)世代の表現型と遺伝子型(各染色体のそれぞれの場所がF0世代のどちら由来かを判別する)の相関性を統計的に解析するQTLマッピング呼ばれる方法で、OBを支配するゲノム領域を探索した。その結果、OBに関与するゲノム領域をc-ski1遺伝子周辺にまで絞り込むことに成功した。しかし、この時点では実際のOBの原因となる遺伝子や性との関係性などが明らかにされていなかった。続いてRobertsらは、先行研究で絞り込まれた領域をさらに精査することで、実はc-skiではなく、その近傍に存在するpax7と呼ばれる転写因子の発現を調節する領域への変異がOBに関与していることを、pax7遺伝子の発現量とOBの表現型との相関性から見出した。そして、このpax7遺伝子の近傍にはマラウィ湖シクリッドにおいてメスを決定づけるW因子が存在することも明らかとなった。つまり、その個体がメスである場合のみに有利に働くOBの原因となる遺伝子の近傍にW因子が飛び込むことで、最終的にメスに特異的なOBが獲得されたと結論づけた。この一連の研究は、OBの形成と性的対立の解消に関するふたつのメカニズムを同時に説明しうる興味深い成果であるが、pax7遺伝子の発現量変化がどのようにしてOBにつながるのか、さらには最も肝心であるW因子の遺伝的実態そのものも明らかにされておらず、今後も精力的な研究が続けられることになるだろう。
⑵エッグダミーを作り出す遺伝メカニズムの解明
上述の通り、haplochromineシクリッドの尻びれにあるエッグダミーは、口内哺育をするために必要な交配行動を誘引する役割をもち、シクリッドの適応放散に大きく貢献したと考えられている。その鍵となる形質ともいえるエッグダミー形成に関わる遺伝子の特定に挑んだのがSalzburgerのグループである。彼らはRNAseq解析を駆使してエッグダミー領域で特異的に発現する遺伝子を網羅的に探索したところ、fhl2b遺伝子の発現が、尻びれの他領域と比較して有意に高いことを発見した。さらに、fhl2b遺伝子の約800bp上流にはSINE(ゲノム中をコピー・ペースト様式で数を増やしていく長さが300bp程度の反復配列で、一般的には役割のないガラクタ配列と考えられている)が挿入していることが明らかとなった。重要なこととして、この遺伝子座へのSINE挿入はhaplochromineシクリッドの共通祖先で起きたため、それ以外のシクリッドではSINEの挿入はなかった。その後に行ったトランスジェニックゼブラフィッシュを用いたレポーターアッセイにより、このSINE配列がエッグスポット領域に存在する虹色色素胞におけるfhl2b遺伝子の発現を担うシス領域である可能性を示した。つまり、haplochromineシクリッドに特徴的なエッグダミーの獲得には、その共通祖先でfhl2b遺伝子の上流に挿入したSINEが引き起こしたということになる。このグループはエッグダミーの獲得という劇的な進化の原因の一端を見出したわけであるが、それは近縁種にもかかわらず表現型が多様なシクリッドを研究モデルにしているからこそ達成できたのだといえよう。また、この研究はシクリッドにおける鍵革新的な形質の進化メカニズムを明らかにしただけでなく、一般的にはガラクタ配列だと考えられてきたレトロポゾンの挿入が関与した可能性を示した興味深い研究でもある。
4.進化の実験場―ビクトリア湖
⑴なぜビクトリア湖のシクリッドを研究するのか?
[サンプルはここまで]
