

昆虫たちの不思議な性の世界
進化するムシたちのラブストーリー
編=大場裕一
B6サイズ/308ページ/3800円+税/2018年6月25日発売/ISBN978-4-910389-03-5(旧ISBN978-4-909383-03-7)
 イグノーベル賞共同受賞の上村佳孝先生、絶賛!
イグノーベル賞共同受賞の上村佳孝先生、絶賛!
何でもアリの昆虫の性を一挙公開!
随所にあるQRコードで動画・音声の視聴ができる!
幼虫が子どもを生む? メスがペニスを持つ? 遺伝子を再利用する?
何でもアリの昆虫の進化の根っこになっているものこそ「性」。あらゆる環境に適応・進化してきた昆虫の「性」をめぐるあらゆるテーマを「プロの虫屋」たちが紹介します!
メスをめぐるオスたちの激しい争い、わが子をまもるための巧みな子育て戦略、オスメスを決定する遺伝子の精巧な働きなど、昆虫たちの面白い行動や分子レベルの仕組みなどを解説していきます。「地球とは昆虫の惑星である」と言われるほど繁栄する昆虫たちの性にまつわるテーマを網羅し、昆虫の魅力、昆虫の持つ性の不思議さ、ひいては生き物にとっての性を、じっくり楽しく感じてもらう内容です。
「昆虫の不思議」がたっぷり詰まった、昆虫ファンなら絶対読みたくなる一冊!
〈3つの特徴〉
1 昆虫の多様性を支える性の営みにフォーカス
書店には昆虫の本はたくさん並んでいますが、こと「性」をフォーカスしたものはないに等しい状況です。交配をめぐるオスたちの激しい争い、わが子をまもるための巧みな子育て戦略、オスメスを決定する遺伝子の精巧な働きなど、「地球とは昆虫の惑星である」と言われるほど繁栄する昆虫たちの性にまつわるテーマを網羅し、昆虫の魅力、性の不思議さを楽しく感じてもらう内容になっています。
2 QRコードによる動画・音声の視聴によって楽しく正確な理解が得られる
文章、写真だけでは伝わりにくい情報を、およそ30箇所の随所に配置されたQRコードを通して紹介しています。雌雄モザイクのノコギリクワガタといった珍しい昆虫から、蛍の幻想的な連続点滅、エンマコオロギの求愛と通常の呼び鳴きとの鳴き声くらべなど、これまで本では伝えられなかった情報を、手軽に提供できる仕組みを持っています。
3 昆虫の「しなやかな性」を見ることを通して、ひいては人間、生き物にとっての性の持つ意味を垣間見る
特に近年、性のイメージが揺らぎ始めていますが、人間でない他の生き物、特に多様性のデパートメントストアと言われる昆虫の、その性のしなやかさの紹介を通して、なぜ性はオスメスの2タイプになっているのか? 性があることの生き物にとっての意味を考え、不思議さを感じられる内容になっています。
【目次】
はじめに|昆虫の性とその意味
まずは〈性〉の基礎知識から/〈性〉はなぜあるのか/〈性〉はDNAエラーを除去する/〈性〉は病原体の進化に対抗する/オスとメスはなぜあるのか/オスとメスが違うことの2つの仮説/働きバチの謎/血縁選択説/昆虫と〈性〉/本書のながれ︱〈性〉は卵から大人まで
第1章 男になるか女になるか|昆虫の性はどうやって決まるのか
性染色体が性を決める?/昆虫の性染色体/多様な性決定要因/遺伝子が性を決める/昆虫には性ホルモンは存在しない/遺伝子の基礎/RNA干渉(RNAi)による遺伝子機能の阻害/性を決めるメカニズム/性決定カスケード/性を決めるメカニズムの多様性/性決定カスケードの進化/コラム オスとメスの見分けかた
第2章 男らしさ女らしさ︱男女の違いができるわけ
幼生生殖/多様な性的二型/体の各パーツにみられる性的二型/性選択/誇張形質のオス・メス差はどのようにつくられるのか/糞虫の角とクワガタムシの大顎/アメンボの触角のオス・メス差はどのようにつくられるのか/擬態のオス・メス差はどのようにつくられるのか コラム 伯方島のヒゲのないヒゲコメツキ
第3章 愛をささやく昆虫たちのことば︱離れていても想いは伝わる
日本人のセンスオブワンダー/昆虫はコミュニケーション戦略のデパートメントストア/鳴く︱音声コミュニケーション/どうやって鳴くのか/ドラミング・タッピング︱振動コミュニケーション/光る︱発光コミュニケーション/匂い︱性フェロモン
第4章 交りの儀︱交配相手をめぐる熾烈な争い
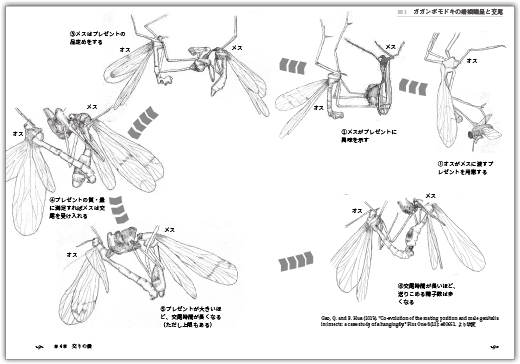
婚姻贈呈/縄張りとスニーキング/交尾様式︱ありえない交尾スタイル/カゲロウのアクロバティックな交尾姿勢/トンボによるハードな交尾姿勢/ゴキブリによる交尾の成功に向けた戦略/カカトアルキによる発展途上(?)の交尾姿勢/カマキリによる命をかけた交尾/ガガンボモドキによる怪奇な交尾姿勢/予備のペニスをもつハサミムシ/生殖器の多様な変化/精子・精包の行方/交尾時間
第5章 処女生殖︱単為生殖とその進化
さまざまな処女生殖/両性生殖と単為生殖の混合繁殖法/コラム 原始的なハチ「カブラハバチ」の産み分け/単為生殖の起源/コラム チビナガヒラタムシの繁殖システム/偽受精交配/コラム オスの遺伝子がのこるとき︱雄性発生する昆虫/一度行ったら引き返せない?︱単為生殖への進化
第6章 男の仕事・女の仕事︱多様な繁殖様式と子育て、そして社会性の進化
子育てをする昆虫/ハサミムシ母親の献身的な子育て/オスが子育てをするコオイムシ類/タガメの父育システムから探るコオイムシ類の父育システムの進化史/父育の進化/社会性昆虫類とカースト制/亜成虫繁殖
-

- 雌雄モザイクのオナシシロオビアゲハとノコギリクワガタ

-
- ガガンボモドキの求愛と交配のようす
-

- シリアゲムシの婚姻贈呈

-

- ホタルのコミュニケーションと警告

【本書で取り上げられている昆虫の一覧】
アカツキシロカゲロウ/アカトンボ/アキマドボタル/ムネクリイロボタル/アサヒナカワトンボ/アザミウマ/アシナガバチ/アトマルキクイムシ/アブラムシ/アメンボ/アワノメイガ/イエバエ/イボトビムシ/イリオモテボタル/ウスバシロチョウ/ウマオイ/ウミホタル/オオアオイトトンボ/オオクジャクヤママユ/オオコオイムシ/オオシロカゲロウ/オオミジンコ/オキシデンタリスカブリダニ/オサムシ/オスへテロ/オナシシロオビアゲハ/オバボタル/カイガラムシ/カイコ/カカトアルキ/ガガンボモドキ/カブトムシ/カブラハバチ/カマアシムシ/カマキリ/カマキリモドキ/カマドウマ/カミキリムシ/カメムシ/ガロアムシ/ガンビエハマダラカ/キイロショウジョウバエ/キジラミ/キノコバエ/キバネシリアゲ/キベリヒゲボタル/キョウソヤドリコバチ/キリギリス/クダマキモドキ/クロバネキノコバエ/クロマドボタル/ケープミツバチ/ケラ/ゲンジボタル/コオイムシ/コオロギ/コガタアカイエカ/ゴキブリ/コクヌストモドキ/コナジラミ/コブハサミムシ/コメツキムシ/ザイノキクイムシ/シオカラトンボ/ジュズヒゲムシ/シュモクバエ/シリアゲムシ/シロアリ/シロアリモドキ/スズムシ/スズメバチ/スペイヤーキノコタマバエ/セイヨウミツバチ/タカサゴシロアリ/タガメ/チチュウカイミバエ/チビカワトンボ/チビナガヒラタムシ/チャタテムシ/ツチハンミョウ/ツマグロオオヨコバイ/ツマグロシリアゲ/ティメマ/トクトク虫/トビイロウンカ/トビケラ/トビムシ/ドラミング/トリカヘチャタテ/ナナフシ/ニイニイゼミ/ニシカワトンボ/ニッポンヒラタキノコバエ/ニュージーランドヒカリキノコバエ/ネジレバネ/ネッタイシマカ/ネバダオオシロアリ/ノコギリクワガタ/ハサミムシ/ハネカクシ/ヒウラカワトンボ/ヒガシカワトンボ/ヒカリコメツキ/ヒゲコメツキ/ヒナバッタ/ヒメボタル/ビワコシロカゲロウ/フタホシコオロギ/ホソクシヒゲムシ/マツムシ/マルトビムシ/ミズスマシ/ミツバチ/ミノムシ/ムカシゼミ/ムネクリイロボタル/メタリフェルホソアカクワガタ/メンガタスズメ/ヤエヤママドボタル/ヤマトシロアリ/ユカタンビワハゴロモ/ワタキバガ
【筆者紹介】
大場裕一(はじめに・第3章):中部大学応用生物学部環境生物科学科准教授。1970年生まれ。北海道大学大学院理学研究科修士課程修了。総合研究大学院大学博士課程修了。博士(理学)。昆虫DNA研究会代表幹事。専門は発光生物学。著書に『恐竜はホタルを見たか』(岩波書店・2016年)、『光るいきもの―海のいきもの』(くもん出版・2015年)、『ホタルの光は、なぞだらけ―光る生き物をめぐる身近な大冒険』(くもん出版・2013年)など、編著に『遺伝子から解き明かす昆虫の不思議な世界』(悠書館・2015年)など。
新美輝幸(第1・2章):基礎生物学研究所進化多様性生物学領域教授。1965年生まれ。名古屋大学大学院農学研究科博士課程(後期課程)修了。博士(農学)。現在、基礎生物学研究所進化発生研究部門/総合研究大学院大学教授。昆虫の翅やカブトムシの角など昆虫が進化の過程で独自に獲得した新奇形質に着目して、昆虫の多様な形質をもたらす分子基盤および進化メカニズムを解明することを目指している。
東城幸治(第4・6章):信州大学大学院理工学系研究科教授。1971年生まれ。筑波大学大学院生物科学研究科博士課程修了。博士(理学)。現在、信州大学学術研究院理学系教授。昆虫類を中心に、クマムシからクマまでの多様な動物の系統地理の研究等に取り組む。著書に『カカトアルキのなぞ』(新日本出版社・2007年)、『ライフサイエンスのための生物学』(培風館・2015年、共著)など。
関根一希(第5・6章):立正大学地球環境科学部環境システム学科助教。1982年生まれ。信州大学大学院総合工学系研究科博士課程修了。博士(理学)。現在、立正大学地球環境科学部助教。高校時に単為生殖へ強く興味を持ち、大学で単為生殖するオオシロカゲロウと出会えた。それ以来、カゲロウなどの水生昆虫類を中心とした繁殖生態や系統地理の研究に取り組んでいる。
