
ここでは、エネルギー資源の開発と利用を通して、人類がいかに人口増加ならびに多様な社会形成を拡大させ、またその過程で自然環境の制約をどのように受けてきたのかについて歴史的に概観する。
1.人類の大移動︱人類学から指摘される正味エネルギーの重要性
人類の生存にとって食物をいかに確保するかは最も重要な事項である。ヒト属(ホモ属)に属する生物種が現れた約200万年前以降を考えると、そのほとんどの期間を狩猟・採集により食物を得ていた。現在においても、カラハリ砂漠にはサン族(あるいはクン族)と呼ばれる狩猟・採集で生計を営んでいる部族が存在する(図1)。このような狩猟・採集の生活は動物の生活様式とほとんど変わることなく、人類学者のリチャード・リーは、サン族の行動をエネルギーの視点から分析したところ、エネルギー収支(狩猟・採集に投入するエネルギーとそれにより得られるエネルギーのバランス)が最も良い状態を無意識に保持していることを明らかにした。リーによれば、そのエネルギー収支は、1キロカロリーのエネルギー投入に対して10キロカロリーのエネルギー獲得を行っており、この場合の正味獲得エネルギーは9キロカロリーとなる。

図1.カラハリ砂漠で狩猟・採集で生計を営むサン族
また狩猟・採集の生活は動物の生態様式と似て自然資源に依存しているが、自然環境が変化した際の反応は人間と動植物では異なることを生態学者のチャールズ・ホールは指摘している。すなわち、自然環境が変化した際に、動植物は遺伝子をベースにして非常にゆっくりとした生物的進化で対応するのに対して、人間はミーム(人間を媒体として増殖する思想的因子)をベースにして生物的進化に比べて短期間で文化的進化をとげることができるとされる。さらに、エネルギーの利用技術の開発は文化的進化の重要な部分であることをホールは指摘している。
約440万年前と推定されるアルディという類人猿が1994年にエチオピアで発見された。人間が直立歩行する際にはチンパンジーに比べて4分の1程度のエネルギー消費であるが、このアルディは直立歩行をしていたもののかなり歩行速度は遅く、さらに木の上にいることが多かったようである。人間の誕生については確定的でない部分が多いが、化石資料に基づくと170万年から180万年前のアフリカであると推定されている。その後、数十万年をかけて、ユーラシア大陸、南北アメリカ大陸、そしてオセアニア大陸へと移動した。前述の生態学者のチャールズ・ホールは、この人類の大移動の過程をエネルギー的視点から以下のように述べている。
狩猟・採集を行っていた人類が最初に開発したエネルギー技術は、石から作った槍先やナイフ(図2)であった。つまり、鋭利性を有するそのような道具はエネルギーを集中させることができるため、より大きな動物をより幅広く狩猟することができる。さらにそれら動物の毛皮を利用することにより寒冷な土地への移住を可能にした。250万年前までには、石を加工していたことが推定される作業場の遺跡がケニヤで発見されている。槍先やナイフを利活用することにより、よりスムーズに動物を獲得する技術、つまりより効率的にエネルギーを獲得する技術を構築していった。次に重要なエネルギー利用技術としては、火の利用である。火を利用することにより調理が可能となり、それまで食物にはなり得なかったものを食物にならしめたし、狩猟・採集生活は当初熱帯地域にのみに限定されていたが火の利用により暖を取ることが可能になり、より寒冷な地域への移住を可能した。こうして、200万年前以降、比較的温暖なアフリカ大陸を離れ、ユーラシア大陸、南北アメリカ大陸、オセアニア大陸に移動していった。

図2.石から作った槍先やナイフ
2.狩猟採集から農耕へ︱エネルギーの流れの大転換
人類の最も重要な文化的進化は農耕の開始である。農耕は約1万年前に肥沃な三日月地帯と呼ばれるチグリス・ユーフラテス渓谷(図3)で起こったとされる。それまでは食物連鎖という制約の中で生きていた人類は、太陽エネルギーを植物の光合成機能を利用することで定期的かつ計画的に獲得することにより、エネルギーの流れを大きく変革することになる。どのようなメカニズムで農耕が始まったのかについては未だ不明な点があるが、進化生物学者のジャレッド・ダイアモンドは、当時の人類が調理場のゴミ捨て場に捨てられた植物の種が発芽したことを発見したことによるのではないかと推察している。さらにダイアモンドは、肥沃な三日月地帯で農耕が開始された理由として、8種類の起源作物がこの地域にすでに自生していたこと、狩猟・採集の生活には不利な条件(水産資源が乏しく、陸上動物も乱獲されたため数が減少していた)であったことを指摘している。肥沃な三日月地帯で農耕が開始されたのは、当該地域の人々が優れていたからという理由ではなく、気候や環境から充分説明できるとダイアモンドは主張する。
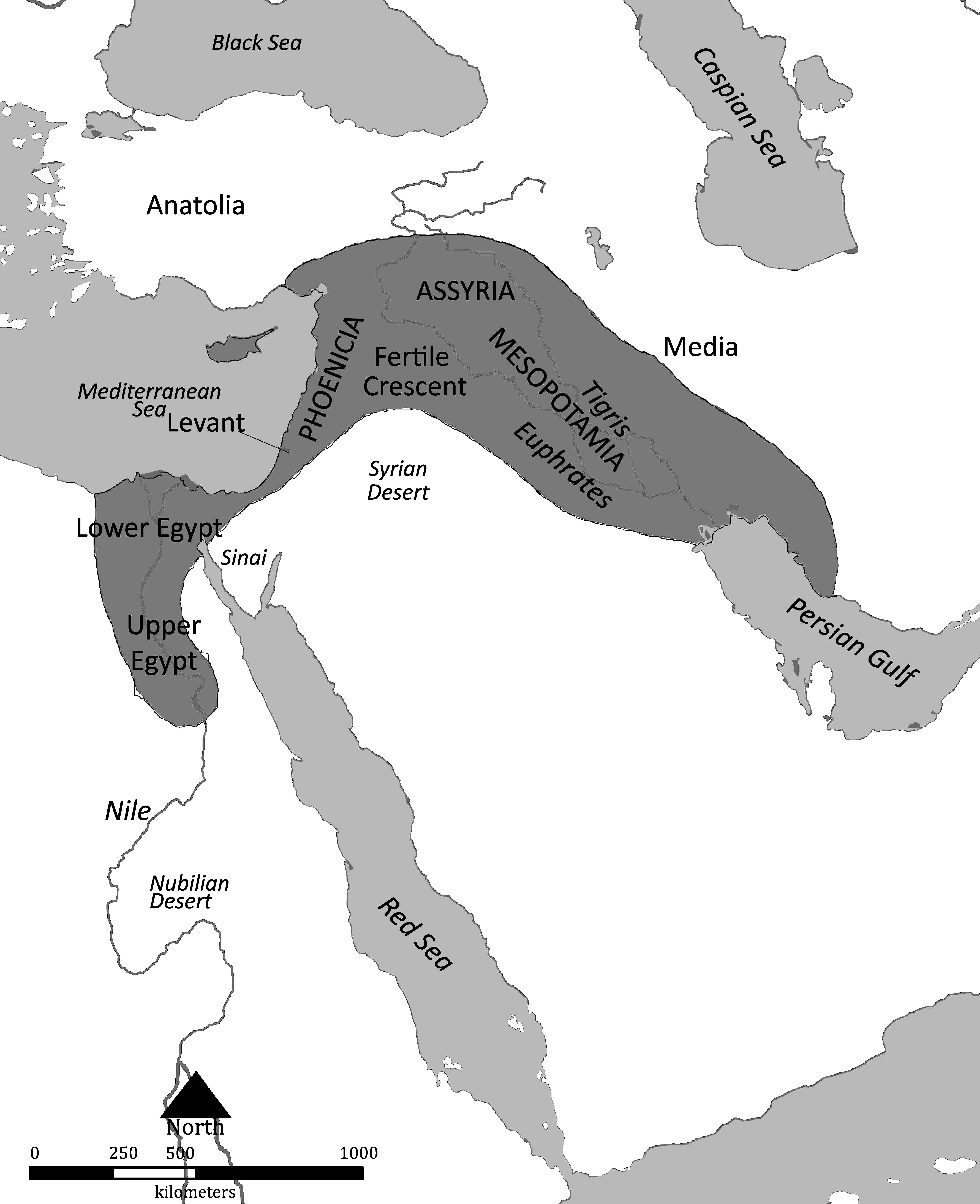
図3.肥沃な三日月地帯
人骨の年代・長さ・強度を調べることによって当時の人類の栄養状態や身体状態を推定することができる。ラリー・エンジェルの調査では、農耕に移行することにより、狩猟・採集をしていた頃と比較して背丈が低くなり、体重が減ったことが明らかにされている。つまり、農耕に移行することにより1人あたりの得られるエネルギーが小さくなり、栄養状態が悪くなったことを意味している。このように農耕は1人あたりのエネルギー獲得の減少につながったが、多くの人に一定のエネルギーを分配することに成功した。すなわち、農耕の導入により、人口を拡大することに成功したと言える。狩猟・採集の時代には、その行為に参加せずに獲得されたものにありつくことはできなかったが、農耕の時代には収穫したものを大量に保存することが可能になるために、農耕という行為をせずとも、収穫物にありつけることができるようになり、そうなることで社会的分化が進んだ。このようなエネルギーの獲得・分配方法の移行により、現在の社会的階層の原型が形成される要因となった。さらに、狩猟・採集の時のように食料を求めて移動する必要がなくなり、ひとつの場所に定住することになり、社会的階層化を促進する効果があったことが推察される。チャールズ・ホールは、このような社会的階層化が市場の原型を形成したことに大きく寄与している点を指摘している。つまり、農耕における収穫率を向上させる知識を持っている人がいる場合、その知識と作物との交換が行われ始め、市場での取引のような行為が行われた。肥沃な三日月地帯で始まった農耕は、その後ユーラシア大陸やアフリカ大陸に伝播していった。
その後、火を利用した金属の精錬技術が発見された。それまでの道具は、石、木材、骨、動物の角などを利用していたが、銅や青銅の道具は非常に丈夫で農耕作業の効率化にも寄与し、その鋭利性のために森林資源を伐採するのに絶大な効果を発揮した。さらに、最も重要な発見として、農耕における家畜の利用と畜産である(図4)。人間と家畜とのコンビネーションは極めて効率的な関係となっている。例えば、穀物の実を人間が食べ、その他の部位を家畜が食べる。また、家畜は農作業や収穫物の輸送にも役立つことができ、糞は肥料となり収穫量の増収につながり、またミルクを提供し最終的にその肉を食べることができる。

図4.紀元前1200年頃のエジプトにおける角牛を利用した耕作
ジャレッド・ダイアモンドは、家畜の重要性について研究を行い、興味深い指摘をしている。そもそも人間にとって重要な家畜になり得る動物は世界に一様に分布していたわけではなく、地理的分布、進化、生態系の3つの要素によっていた。しかし最も面積が大きく幅広い地理的特性と多様な生態系を有していたユーラシア大陸で、家畜化に成功した動物が多かった。人間は様々な動物を家畜化し利便性を享受しようと試みたが、実際に家畜化に成功した動物はわずか14種類(羊、山羊、牛、豚、馬、ヒトコブラクダ、フタコブラクダ、ラマ/アルパカ、ロバ、トナカイ、水牛、ヤク、バリ牛、ガヤル)であり、そのうちの13種類はユーラシア大陸で家畜化に成功し、他の大陸に伝播された。多くの動物が家畜化されなかった理由として、餌の経済的効率が低い点、成長が遅く経済的効率が低い点、捕獲状態での繁殖活動が不活発な点、気性が荒かったりパニックになりやすい点、集団行動に適さない点などがダイアモンドにより指摘されている。最終的に家畜化に成功すると、農作業の効率が向上するばかりでなく、生産物の輸送を可能にすることで交易を促進し、さらには市場を形成することにおいても優位性を発揮することになった。このような家畜導入のプロセスは、結果として人間社会の発展程度の地域性を生じることにつながった。実際、狩猟・採集の時代の遺跡に比べて農耕時代の遺跡の方が多様性に富んでいることがわかっている。ダイアモンドが家畜の研究を行うに至った経緯は、彼が鳥類の研究でパプアニューギニアを訪れた際に、島の若者から「何故白人はたくさんの文明的なモノを作りだせるのに我々パプアニューギニア人は作り出せないのか?」との素朴な質問がきっかけであった。ダイアモンドは詳細な調査を積み重ね、ひとつの、そして素朴な結論にたどり着いた。パプアニューギニアには有用な家畜の利用がなされなかったため、人間は食物を得るために四六時中働く必要があり、文明を進化させる時間がなかったという結論だった。すなわち、文明を進展させるための効率的なエネルギー利用に成功しなかったのである。逆に家畜の利用に成功すると、太陽エネルギーのより効率的な利用が可能となり、人口を増やし社会を発展させることができた。以上のことをエネルギー資源論の観点からみると、食料を集中的に生産し貯蔵することでエネルギーの余剰を生み、このことが人間の活動を本質的に変化させ、人間の文化的進化を促進したことをチャールズ・ホールは指摘している。
3.エネルギー利用の進化と人間の文化的進化
これまで見てきたように、人類はエネルギー利用を拡大することにより、さらなるエネルギー資源獲得のための能力を身につけていった。フレッド・コットレルは、エネルギー利用の進化と人間の文化的進化の関係の重要性について述べている。とりわけ、余剰エネルギー(獲得されたエネルギーから獲得するのに直接・間接的に使用したエネルギーを差し引いたもので、正味エネルギーと同等の意味)が重要な役割を果たしてきたことを指摘している。
余剰エネルギーの概念を理解するために、コットレルはいくつかの例を用いて説明している。穀物の場合の例では、以下のようになる。もし収穫した穀物から得られる種子の量がその収穫を得るために蒔いた種子の量より少なければ、次回の収穫は今回の収穫より少なくなる。その一方で、前回蒔いた種子の量を十分賄えるだけの収穫があれば、次回の収穫は今回の収穫より多くなり、多くの余剰エネルギーを獲得することができ、その余剰分で収穫のための労働力のためのエネルギーばかりではなく、道具や機械を作るためのエネルギーとしても広範に使用することができる。また、別の例では、道沿いの木にブラックベリーが実っており、それらの実は人が少し手を伸ばせば採取できる位置にある場合である。散歩している人が、これらの実を取って食べる際に、手を伸ばしてブラックベリーを採り口に運ぶエネルギーは、ブラックベリーを食べて得られるエネルギーより小さいため、余剰エネルギーを得ることができる。その一方で、草原の中の野ウサギを素手で捕まえて食べようとする人は、捕まえるために使用するエネルギーが、捕まえて食べることにより得られるエネルギーより大きくなってしまうため、余剰エネルギーを得ることはできない。
現代の複雑な社会においては、すべてのモノのやり取りは極めて複雑ではあるが、本質的な点では上記の例で述べたような余剰エネルギーの原理で考えることができる。つまり、ある社会において生産するエネルギーより消費するエネルギーが大きい状況を継続すれば、いずれその社会全体がエネルギー不足に陥る。この場合、別の余剰エネルギーを有する社会からエネルギーを提供される必要がある。古代の社会においては、頻繁に社会全体が余剰エネルギー不足により社会全体が崩壊していた。逆に、豊かな余剰エネルギーを獲得できた社会には繁栄がもたらされた。
以上のように、社会が経済の拡大化や多様化を達成するためには、より大きな余剰エネルギーを有することの必要性を初めて指摘したのはコットレルであった。人間の文化的進化や物質的な生活水準の向上の歴史は、より大きな余剰エネルギーを生み出すエネルギー資源へ如何にアクセスし開発するかに直接的に関連していた。植物、バイオマス、家畜などの生命由来のエネルギーから、風力や水力、さらに化石燃料そして電力へと、使用するエネルギー資源を変遷させることは、エネルギー余剰を拡大することであり、結果として1人あたりの生産性を向上させてきた。高い余剰性を有するエネルギーは、エネルギー獲得に要するエネルギーが少ないため、その余剰分を他の経済活動に振り分けることができる。その結果、経済の拡大化と多様化を達成することができる。余剰エネルギーの創出と分配は、特に19世紀から20世紀に起こった間欠的で急速な成長を理解する要因であると、その重要性をコットレルは指摘している。さらに、静的な量としての余剰エネルギーばかりでなく、生態系を含めた動的なエネルギーフローにより、如何に余剰エネルギーを有効利用しているかを、物理学的アプローチによって分析することの重要性を指摘している。
4.繰り返された古代帝国の崩壊︱エネルギー的視点から
前述したように、人類は農耕を開始することにより余剰作物(すなわち余剰エネルギー)を広く分配することが可能となり、結果として社会的分化が進み階層構造が形成された。その後、時代を重ねるに従って貧富の格差が生まれ、富と権力の集中が起こり、軍事的体制は次第に組織化・巨大化し、結果として、一定の地理的なエリアを統治する帝国が生じた。
考古学者で歴史学者のジョセフ・テインターは24の古代帝国の栄枯盛衰を調べ、共通のパターンがあることを指摘している。家畜を伴った農耕、鉱山開発と精錬技術による金属の利用は余剰エネルギーの生産を促進させ、結果として経済活性を促進し、ひとつの都市の人口と経済規模を拡大させる。そのような経済発展を基盤にして、市場や社会インフラを整備することや豊富な食料を確保することにより繁栄を築くと、そのような繁栄は周辺の都市の人々をも魅了し、その繁栄した都市に取り込まれていく。取り込まれた人々は、農耕作業に従事したり、金属生産・加工を行うなどして、余剰エネルギーの生産に貢献することにより、その都市は成長を繰り返すことになる。しかし、その一方でその都市が地理的に拡大を続けていくと、その中央への穀物提供(税金の役割)のための運搬エネルギーが段々と嵩むことになり、本来の歳入が運搬エネルギーに消えてしまうことにつながり、結果としてそれまでの成長に抑制力が働く。結果として何かのタイミングでひとつの重要な歯車が狂うと、全体のバランスが失われてしまう。
歴史学者のジョン・パーリンは著書『森と文明』の中で、多くの古代文明の崩壊はエネルギー供給源である森林資源の喪失であることを指摘している。前述したように、火を利用した金属の精錬技術の発見は、効率的に森林資源を伐採するのに絶大な効果を発揮した。つまり、エネルギー獲得速度が石器に比べて格段に向上したのである。大量の森林資源を燃焼させることにより銅などの鉱石を大量に精錬することが可能となり、得られた金属の交易を通して富を獲得することができた。さらには、森林を伐採した後の土地では農業が可能となるため、農作物の生産・交易によっても生活の豊かさを向上することができた。このような森林資源に根ざした良好な状況は1世紀ほど継続するが、肥沃だった森林土壌が農耕で使用され続けると、だんだんと肥沃さが失われ、結果として作物の収量が減少する。それまで成長を続けてきた文明は、鉱石の品質低下と作物の収量低下により衰退を始める。以下では、古代帝国の衰退をエネルギーの視点から分析した例を紹介する。

図5.古代ギリシアのポリスの象徴であったアテネのアクロポリス
紀元前8世紀頃の古代ギリシャでは、多くの国家的な機能をもった小規模な都市国家(ポリス)が成立していた(図5)。紀元前5世紀前半にアケメネス朝ペルシアとアテネ(現代の民主政の源流を確立したと言われる)を中心とするポリス諸都市との間でペルシア戦争が起こり、最終的にはギリシャ側が勝利した。ジョン・パーリンは、このようなギリシャ側の勝利の要因として、アテネ周辺の丘に存在した豊富な森林資源を指摘している。ペルシア戦争における歴史的海戦であるサラミスの海戦においては、多数の軍船の建造が必要だったが、豊富な森林資源がそれらの建造を支持した。その一方で、パルテノン宮殿の建設などの都市における資材・エネルギー需要の拡大や、ラウリオン鉱山から産出される鉱石の製錬(精錬された銀は武器にも使用される)におけるエネルギー需要の拡大が続くと、森林資源が重要な戦略物質である認識もされていった。その後、アテネとスパルタがギリシャを二分して覇権を争ったペロポネソスの戦い(紀元前431年から404年)の頃には、森林資源や農耕土壌の衰退が顕在化しており、森林資源の確保が勝敗を左右する要因になった。スパルタ側は昔の敵だったペルシアと同盟を組むことによりペルシア側の森林資源の利用が可能となり、結果としてスパルタが勝利したが、社会を支える森林資源というエネルギー基盤の喪失からギリシャ全体の衰退を招くことになった。古代ローマ帝国は、イタリア半島中部に位置した多部族からなる都市国家を起源として、国力が衰弱したギリシャの都市国家を次々に征服し、領土を拡大していった。古代ギリシャがポリスの連合体で形成されているのに対して、古代ローマは中央政権が支配する領土国家であった。2世紀頃になると、ローマ帝国の規模は最大となり、地中海世界の全域を支配するまでになった(図6)。この時、帝国は100万人を超える住民を抱えており、征服・公共事業などの様々な活動は主として人力エネルギーに依存していた。従って、そのような人力エネルギーを支えるための食料供給の土地の確保が重要であった。ローマ帝国のエジプトへの侵攻と服従は、カエサルのクレオパトラへの情欲ではなく、ローマ人によって劣化させられたイタリアの農耕土壌を背景とした、食料供給のための土地確保であったことを、チャールズ・ホールは指摘している。「エジプトはナイルの賜物」の言葉の通り、ナイル川の定期的な氾濫によって形成された肥沃な土壌の恵みで、エジプトの壮大な文明が築かれた。このようにして、商業・貿易は栄え、公共事業等の土木・建築技術も進展し、2世紀頃には人々にとって最高の時代を迎えていた。一方で、自然資源(穀物、森林資源、太陽エネルギー)の変動に影響を受ける経済の浮き沈みという問題が存あった。つまり、ローマ人は自然から与えられるエネルギー供給能力を超えるエネルギー消費をしていたことを意味する。結果として、生きるために必要なエネルギーを供給していた周辺の自然資源の劣化が始まり、より遠くの自然資源に依存するようになった。そうなると今度は輸送エネルギーが発生するため、正味の生産性は落ちることになる。このような状況に対応するために、政府は金や銀などの当時の貨幣の改鋳を行った。しかし、この貨幣改鋳により過度なインフレが起こり、交易における購買力は低下した。
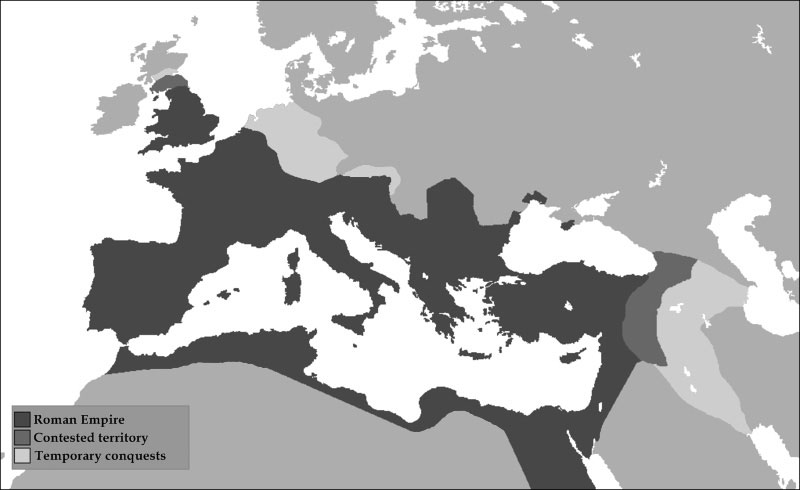
図6.古代ローマ帝国の最大勢力図
ジョセフ・テインターはローマ帝国の崩壊をエネルギーのバランスの崩れで説明している。ローマ人の自然資源の獲得の方法は侵略・征服であった。ある領土に形成された自然資源は、太陽エネルギーを蓄積したものである。そして、鉱物資源は人間のエネルギーで採掘し、森林資源を使用して精錬しなくてはならない。このような仕事は骨が折れるため、ローマ人はその種の作業には手を出さず、それらを有する地域を征服してそれらを獲得する手段を選んだ。被征服国からエネルギーや富の流れができることによってローマ帝国が拡大していくと、輸送のためのエネルギーが増すとともに、帝国全体を統治する仕組みが複雑性を増してくる(例えば食料を適切に生産し適切に分配する仕組みは規模が大きくなるほど難しくなる)。そのため、統治のためのエネルギーも増してくる。このような領域の拡大と複雑性の増加に対応するための余分なエネルギー消費が増えることになる。最終的に臨界点を超えると、全体としての劣化が始まる。5.産業革命︱低エネルギーから高エネルギー社会への転換
経済学的観点から見ると、富の源泉は、土地→労働→資本と変遷してきた。英国の経済史学者であるエドワード・アンソニー・リグリー(1931年〜)は、著書『エネルギーと英国の産業革命』の中で、労働と資本については原理的に無限に拡大が可能であるが、土地は有限性であることを指摘した。ここでは、土地は食料生産の場のみならず、金属精錬などに使用される木材資源の場でもある。例えば、金属の生産量を増加させようとすると、多くの森林資源から木材を調達することになる。最初は優良な森林資源を利用できるが、次第に劣悪な森林資源を利用することになり、最終的に金属の生産量は頭打ちとなる。従って、産業革命以前においては土地の制約に強く依存した社会であり、リグリーはこうした社会を有機経済と呼んだ。有機経済とは、すなわち太陽エネルギーによる光合成を介して成立する経済である(太陽エネルギーの他に風力や水力エネルギーもあるがこれらの量は相対的に小さい)。
しかしながら、光合成によるエネルギー変換効率はさほど大きいものではなく、地球に降り注ぐ太陽エネルギーの約0・1〜0・4%程度と言われてる。生態系の食物連鎖を考えても光合成は最も基盤的な部分であり、人力や家畜による力学エネルギーを生み出すばかりでなく、森林資源の燃焼による熱エネルギーを生み出すことができる。森林資源の燃焼による熱エネルギーは多くの産業過程で不可欠なもので、ガラス工場、煉瓦作成、ビール醸造、織物染め、金属精錬に使用された。その意味で、有機経済においては森林資源が熱エネルギーを生み出せる唯一のエネルギー源であったが、その森林資源の生産能力は4000㎡あたり乾燥木材として年間約1〜2トンほどであった。ちなみに2トンあたりの乾燥木材のエネルギーは1トンあたりの石炭のエネルギーに相当する。1トンあたりのエネルギー量にそれほど大きな違いはないが、森林資源は年間に生産される量が、光合成により得られるエネルギーに制限を受けることになる。交易を通してこのような制限から多少緩和されることはあるが、どんなに経済状況が良い状態であっても、根本的にその制限から逃れられることはできないことをリグリーは指摘している。
リグリーは、有機経済社会において、人力に対する家畜のエネルギーの比率の重要性を指摘している。多くの社会において、その比率が人間の栄養摂取状態よりも1人当たりのエネルギー消費量を決定する要因であると述べている。例えば、農耕作業において馬は人間の6倍程度の仕事を実行することが可能であり、多くの馬や雄牛を有していれば農耕作業の効率性が飛躍的に向上し、人力エネルギーの大幅な削減を達成することができる。さらには、家畜のために牧草地を確保することは、人間の食料を生産するための土地を減少させることになり、結果として人口増加は抑制され、1人あたりの農業生産量を向上させる。さらに、リグリーは輸送における家畜エネルギーの利用の重要性も指摘し、この家畜エネルギーが産業や鉱山採掘の活動における1人あたりの生産量を向上させたことを指摘している。結果として、家畜エネルギーの有効利用が有機経済における生活水準を大きく左右した。
有機経済において、人々は光合成が生み出すエネルギーの制約のもとで生きる必要があった。経済成長は極めてゆっくりとしたもので、人間が作り出す政治・経済・社会の不完全さとは関係なく、継続的な成長は実現されなかった。このような有機経済の成長限界性については、アダム・スミスやデヴィッド・リカードも指摘していた。土地の生産性が経済成長の決定因子であり、生産性の低い土地に対する投資の利益率は低いものであった。土地の生産性が高い間は、経済は成長し労働者の給料も上がるが、そのような好条件の土地が減少していき、生産性の低い土地に資本と労働が投入される段階になると、そのような経済成長は続くことができない。
これらの指摘に対して、マルサスは著書『人口論』の中でさらなる考察を行った。経済成長に伴う給料の向上は、生活水準の向上により死亡率を下げ、早期婚姻を促すことにより人口増加率を向上させる。経済劣化の影響を受けない人口は等比級数的(掛け算的)に増加する一方で、土地の制約に起因する物質的な生産は等差級数的(足し算的)にしか増加しない。等比級数的なものは等差級数的なものを凌駕してしまうため、労働者に困窮をもたらしてしまい、結果として人口増加は抑制される。すなわち、どんなに技術改善が行われたとしても自然が提供する制約からは逃れることはできないと認識されていた。人間の全ての活動はエネルギーに関連しており、自然から提供されるエネルギーの総量で人間の活動は制約を受けるという単純な構造であったのが有機経済の根本である。
このような有機経済におけるエネルギー制約から人間社会を解放したのが、石炭資源であった。石炭資源の利用は英国で先導的に進められ、まずは18世紀初頭まで熱エネルギー供給における木材資源の代替とされた。その後、蒸気機関が発明されると石炭資源により生産される熱エネルギーは力学的エネルギーに変換することに成功した。こうなると石炭資源の利用は社会の様々な場面で有機経済における制約から人間社会を解放していった。このようなエネルギー革命により、エネルギー使用量は石炭資源以前と比較して240倍以上となった(図7)。産業革命がどのように始まり、どのように社会の生産性を拡大し、多くの人々を貧困から救ったのかについては多くの書物で述べられているが、そのような急速な発展を可能にした最大要因がエネルギーであったことが述べられることは少ない。

図7.19世紀のイングランド北西部マンチェスターの工場群。技術進歩によって石炭は産業革命の主要なエネルギー源となり、その消費量は大幅に増加した。
リグリーは、英国が有機経済から安価で豊富な石炭エネルギーに基づいた経済に転換した状況を、以下のように述べている。英国は、16世紀から17世紀頃にかけては、他のヨーロッパ諸国と比較して相対的に劣るような経済・生活レベルであった。チューダー朝時代(1485〜1603年)においては新規の鉱山事業や新興産業を企てる動きも見られたが、多くは他のヨーロッパ諸国の専門家頼みの状態であった。金融事業に関しても特段の優位性はなく、国際貿易においては、単純な工場制手工業による羊毛製品の輸出が卓越していたのみであった。農業技術の進歩については、特に北海沿岸の低地帯諸国(現在のベルギー・オランダ・ルクセンブルクが占める地域)から新技術を導入する状態であった。さらには、政治的影響力や軍事力についても、当時のスペインやフランスにも及んでいなかった。そのような状況が19世紀になると大きく一変することになった。ロンドン万国博覧会 (1851年)の時代には、英国はヨーロッパで最も発展した国となっていた。そして、ロンドンは世界で最も影響力のある商業都市となり、国際貿易の拠点となった。蒸気機関の導入により、産業の幅が格段に拡大し輸送能力も飛躍的に向上した。このように、安くて豊富な石炭エネルギーへの転換とその利用技術である蒸気機関の導入が英国に大きな転換をもたらしたが、その中でも農業と輸送の向上が重要であったことをリグリーは指摘している。農業については、穀物の収穫量が16世紀に比べて19世紀は約2倍となった。農耕や畜産の拡大は人口増加による食料需要ばかりでなく第2次産業の素原材料の需要も拡大していた。そして、第1次と第2次産業で生産されるものの拡大は輸送需要の拡大をもたらした。アダム・スミスが提唱した分業の概念も成長過程を加速させた。スミスの著書『国富論』ではピンの製造を分業することにより生産数を飛躍的に増加させることができる(分業せずに1人で作成すると20個も生産できないが、分業すると1人あたり4800個生産できる)と述べているが、輸送手段・インフラの向上により生産効率はさらなる飛躍が可能となる。さらに、都市と都市が有機的に結合されると、それぞれの都市は活性度を帯び、人口増加へと繋がった。以上のように、真のエネルギー革命は波及効果が広く大きく、各要素が有機的に結合し相乗効果を発揮していった。
6.ジェボンズのパラドックス
石炭の導入によるエネルギー革命が社会に及ぼした影響の大きさについては前述した。低価格で豊富な石炭エネルギーが社会に行き渡り、社会全体のエネルギー効率も向上したにもかかわらず、人間はより多くのエネルギーを消費する革新的な方法を次々に発見し、結果としてエネルギー消費量を増やしてしまうというパラドックスを、前述した英国の経済学者ウィリアム・ジェボンズ(1835〜1882年)(図8)が1865年に著書『石炭問題』で指摘している。彼の名にちなんで、このパラドックスは「ジェボンズのパラドックス」と呼ばれる(経済学的にはリバウンド効果とも呼ばれる)。ジェボンズはこのパラドックスを英国の製鉄産業を例にして説明している。技術革新により、少ない石炭で鉄を生産できる溶鉱炉を開発すると利益が上がるので、鉄をさらに生産するための新規投資が行われ、鉄の値段は下がり新たな需要を喚起する。結果として、省エネルギー技術は消費エネルギー全体を減らすことに貢献せず、むしろ総消費エネルギーを増やすことになる。

図8.ウィリアム・ジェボンズ
ジェボンズは1835年に英国リバプールで生まれ、ロンドン大学で学位を取得した後は大学の講師をしながら、金の市場に関する書籍を出版していた。そして1865年に出版された『石炭問題』で名をはせることになる。そして、1871年には『政治経済の理論』と題した書籍を出版し、現在でも数理経済学の重要な基本教科書のひとつとされている。その後、1882年には英国海峡を水泳中に46歳の若さで溺死している。20世紀における経済学者の代表的存在であるジョン・メイナード・ケインズ(1883〜1946)はジェボンズのことを、19世紀における偉人の1人であると述べている。さて、このジェボンズのパラドックスに関しては、現在においても様々な角度から研究され多くの学術論文で発表され続けている。このパラドックスは、成長盛りの社会においては成立するが、近年の先進国のような低成長の社会には当てはまらなくなってきたことも指摘されているが、実際問題として先進国において省エネ技術を導入してもエネルギー消費量や二酸化炭素排出量を減らせないため、このパラドックスを簡単に棄却することができない位置付けにもなっている。
状況はつまりこういうことであろう。よく知られているように、日本は省エネルギー技術の優れた国である。1970年代の2回の石油ショックという負の状況を糧に、正の状況に転換した良好事例と言える。日本人が本来有する節約志向と手先の器用さのコラボレーションの産物と言えるだろう。しかし、このような省エネルギー技術に優れた国が、エネルギー消費量を総体として減らせていないのである。例えば、ある家庭でガソリン車をハイブリッド車に買い換えたとすると、毎月のガソリン代が安く済むことになり、ガソリン代で浮いたお金で新規に家電製品を買い、結局エネルギー消費の軽減につながらない。
エネルギー効率の向上は様々なレベル、すなわち資源採掘から工場での生産活動を経て生産物の輸送、そして最終的な使用に至るまでのコスト削減に貢献するが、そのコスト削減がさらなる消費を刺激してしまう。エネルギー効率向上の各要素の効果を読み取ることは決して難しくはないが、全体としての波及効果を分析するためには、それぞれの各要素の相互作用を体系的に理解することは極めて複雑となる。
その一方で、このようなジェボンズのパラドックスの効果は近年では影響が少なくなってきているとの指摘もある。つまり、効率的な機械の導入によりエネルギーの使用と経済活動との関係性が弱くなっている(デカップリング現象とも呼ばれる)というものである。ザ・ニューヨーカー誌のデヴィッド・オーウェンは、デカップリング現象によってジェボンズ効果が減少する傾向に対して異議をとなえている。例えば、人件費やエネルギー費用の安い国に工場を移動させる時に、本来自国で使用するエネルギーを他国に移動させる場合を考慮していないことを指摘している。さらにデンマークの研究者であるジョルゲン・ノルガードは、デカップリング現象が主張する、経済におけるエネルギーの役割の縮小は本質的な誤認であることを指摘し、エネルギー効率向上によるエネルギー生産の経済規模全体に占める割合は確かに減少してきているが、逆にその小さい割合で経済全体を支えていると言え、重要性は増していると主張する。
生態経済学者のブレーク・アルコットはジェボンズのパラドックスの現代における大いなる存在について、以下のように述べている。もし、エネルギー効率が向上し、全体としてのエネルギー使用量を削減できるのであれば、エネルギー効率を悪くすればエネルギー使用量は増加するはずである。しかし、実際にはそのようになることはなく、例えば1800年頃のジェームズ・ワットの時代においては、現代の技術よりエネルギー効率が低かったが、この技術レベルが現代までそのままのレベルで維持された場合、現代より多くのエネルギーを使用することはないことは想像に難くない。つまり、我々はエネルギーへの依存性を減少させている(デカップリング現象)のではなく、エネルギーをテコの原理を利用して上手に使用する方法を発見しているに過ぎないのである。
1984年から2005年の間に、米国の電気製品の生産量は約66%増加した。増加要因のひとつは人口増加が挙げられるが、1人あたりのエネルギー消費量は上昇し、単位GDPを生むのに使用するエネルギー量は半分程度になった。人口増加自体もジェボンズ効果であり、エネルギー効率が向上すると多くの人口を養うことができ、結果として多くのエネルギーを消費する。このようにジェボンズのパラドックスの本質は、エネルギーを節約してもその節約されたエネルギーで別の経済行為を行ってしまうということであり、これを避けるためにはエネルギー節約分で浮いたお金を使用しないことであるが、これは現実的ではない。結局のところ、人間の歴史を通してジェボンズのパラドックスは日常的に起こっている現象であり、その矛盾から逃避できないことを示唆している。
ジェボンズは1882年に事故で溺死してしまったため、エネルギー源としての石油や天然ガスの存在を知り得ることはなく、石炭の時代を生きた。彼は石炭の利便性と生産性の高さを熟知していたため、風力・水力・バイオ燃料(当時は木材や鯨油)は石炭の代替にはなり得ないことを主張していた。彼の著書『石炭問題』の最後において、英国が直面している問題というのは、短期間に豪華な生活を送るのか、より長く続く普通の生活かの二者択一であることを指摘し、彼自身は後者(現在で言うところのサステナブル社会)を好むと述べている。ジェボンズのパラドックスは時代を超えて現在でも中心的な存在となっている点をデヴィッド・オーウェンは指摘している。
7.人口変動とエネルギー
人口は微視的に見れば常に増減を繰り返してきたが、巨視的に見れば増加をしており、人口学者エドワード・ディーヴェイ(1914〜1988)は急激に人口が増加したタイミングとして、道具作成を始めた時、農耕を始めた時、そして産業革命の時の3時点を指摘している。この3時点はいずれもエネルギーと密接に関連している。前述したように、人類史上初のエネルギー技術である槍先やナイフ等の道具を利活用することにより、熱帯地域にのみに限定されていた狩猟・採集生活は、より寒冷な地域への移住を可能した。さらに、農耕は1人あたりのエネルギーの獲得の減少につながったが、逆に多くの人に一定のエネルギーを分配すること、すなわち人口を拡大することに成功した。そして、産業革命の時には石炭資源へのエネルギー革命が起こり、自然から提供されるエネルギーの総量で人間の活動が制約を受けることがなくなった。
人間がまだ原始的な生活をしている時には、出生力、死亡力、移動の3つの要素で人口の増減を議論することができるが、人間の社会が高度化してくると、経済的あるいは社会的な要因も影響を及ぼし複雑性が増してくる。とりわけ、エネルギー資源論的な観点からは、人口増加と食料獲得手段の進化との関係が重要である。このような関係は人類学者によって検討され、人口の規模は食料獲得手段の技術、すなわち自然環境から効率よく恵みを得る技術の改善と強い関連があることがわかっている。そして、そのような関係は生産性が再生産性(当初の規模を維持できる能力)を決定づけるということに帰結される。前述したように、有機経済において生産される全てのものは光合成により生み出されるため、エネルギー制約を受ける。リグリーは再生産性の重要性について指摘している。生産性ばかりでなく再生産性によって経済状況が影響を受けることはマルサスも指摘している。リグリーは有機経済における拡大範囲の差異として、18世紀のオランダとフィンランドとの差異の例を述べている。オランダでは農業、産業、貿易・輸送、その他の比率が、41対32対16対11に対して、フィンランドのそれは80対4対1対15であった。すなわち、フィンランドと比較してオランダでは自然から効率良く恵みを享受することにより有機経済をより良く発展させた。このような大きな差異は、経済成長がエネルギー制限を受けてしまう有機経済における生産と再生産との相互作用の重要性を示している。
